|
|
CURSO DE BIOLOGÍAAlejandro Porto Andión |
|
|
|
|
|
|
|
Inicio Temas de Células Aula virtual |
|
|
|
|
![]()
TEMA 15: BIOENERGÉTICA.
1.-CONCEPTO DE METABOLISMO.
Se denomina metabolismo (o también metabolismo intermediario) al conjunto de reacciones químicas enzimáticamente catalizadas que tienen lugar en la célula. Esta definición, si bien es correcta, resulta un tanto incompleta, ya que no da idea de que el metabolismo no es un mero conjunto de reacciones, sino una actividad química altamente ordenada y llena de sentido cuyo objetivo es la correcta manipulación de la materia y la energía por parte de la célula para así mantener el estado vital. En sucesivos apartados de este tema iremos profundizando más en la verdadera naturaleza y significado del metabolismo.
2.-FUENTES DE MATERIA Y ENERGÍA PARA EL METABOLISMO.
La maquinaria de transformación energética de las células está formada por biomoléculas orgánicas. Estas biomoléculas poseen características similares en todas las formas de vida. Sin embargo, existen grandes diferencias entre distintos tipos de células en lo que se refiere a la forma en que obtienen de su entorno el carbono que necesitan para construir los esqueletos de sus biomoléculas constituyentes, así como otros elementos, como el nitrógeno y el azufre, que necesitan incorporar a algunas de ellas. Atendiendo a este criterio podemos distinguir dos tipos de células:
a) Células autótrofas (también llamadas litótrofas).- Obtienen el carbono en forma de CO2 y otros elementos como el nitrógeno y el azufre en forma de sales minerales (nitratos y sulfatos), es decir, toman la materia de su entorno en forma de materia inorgánica y son capaces de transformarla después en materia orgánica. La palabra "autótrofa" significa etimológicamente "que se alimenta por sí misma" aunque quizás sea más adecuada la denominación "litótrofa" ("que se alimenta de piedra") si nos tomamos la licencia poética de llamar "piedra" a la materia inorgánica que estas células toman de su entorno. Las células autótrofas son relativamente autosuficientes ya que no dependen de otras células para alimentarse.
b) Células heterótrofas (también llamadas organótrofas).- No pueden utilizar el CO2 ni las sales minerales, es decir, la materia inorgánica, y por lo tanto deben obtener tanto el carbono como otros elementos en forma de sustancias orgánicas, tales como monosacáridos, aminoácidos, etc., que han sido elaboradas previamente por las células autótrofas, de las cuales dependen para su alimentación. La palabra "heterótrofa" significa etimológicamente "que se alimenta de otros".
Por otro lado, aunque todas las células transforman la energía que extraen de su entorno en energía química de los enlaces de sus biomoléculas constituyentes, existen grandes diferencias entre distintos tipos de células en lo que se refiere a la forma en la que obtienen dicha energía. Atendiendo a este segundo criterio también podemos dividir las células en dos grandes grupos:
a) Células fotótrofas ("que se alimentan de la luz").- Obtienen la energía que precisan en forma de energía radiante asociada a las radiaciones electromagnéticas, fundamentalmente la luz visible.
b) Células quimiótrofas.- Obtienen la energía que precisan a partir de reacciones químicas exergónicas, concretamente reacciones redox, en las que determinadas sustancias ceden sus electrones (se oxidan) a otras que tienen tendencia a aceptarlos (reduciéndose así), lo cual conlleva un desprendimiento de energía. Estas células pueden a su vez subdividirse en aerobias, si utilizan el O2 como aceptor último de electrones en sus reacciones redox, y anaerobias, si utilizan alguna otra sustancia, generalmente de naturaleza orgánica. Muchas células pueden funcionar de modo aeróbico si hay oxígeno disponible y en modo anaeróbico en caso contrario; se dice que son facultativas. También hay células quimiótrofas que en ningún caso pueden utilizar el oxígeno e incluso resultan intoxicadas por él; se dice que son anaerobias estrictas.
Teniendo en cuenta simultáneamente los dos criterios enunciados podemos clasificar las células vivas en cuatro grandes grupos según sean las fuentes de materia y energía que utilizan para su metabolismo:
|
TIPO DE CÉLULA |
FUENTE DE MATERIA |
FUENTE DE ENERGÍA |
|
Fotolitótrofas |
Materia inorgánica |
Luz |
|
Fotoorganótrofas |
Materia orgánica |
Luz |
|
Quimiolitótrofas |
Materia inorgánica |
Reacciones redox |
|
Quimioorganótrofas |
Materia orgánica |
Reacciones redox |
Es conveniente reflexionar sobre el hecho de que las células quimiolitótrofas y quimioorganótrofas utilizan respectivamente la materia inorgánica y la materia orgánica no sólo como materias primas para las construcción de sus biomoléculas sino también como sustancias dadoras de electrones en las reacciones redox mediante las cuales obtienen su energía. El mismo doble papel desempeñan respectivamente la materia inorgánica y la materia orgánica en las células fotolitótrofas y fotoorganótrofas, ya que, como veremos más adelante, éstas células, en realidad, también obtienen su energía a partir de reacciones redox, las cuales, a diferencia de las que tienen lugar en las células quimiótrofas, son endergónicas, por lo que requieren un aporte energético en forma de luz.
La mayor parte de las células vivas son o bien fotolitótrofas (células verdes de las plantas superiores, algas, cianofíceas y bacterias fotosintéticas) o bien quimioorganótrofas (células animales, células de los hongos y la mayor parte de los microorganismos). Sin embargo existe un reducido grupo de microorganismos quimiolitótrofos y fotoorganótrofos que no deben ser considerados meras anécdotas de la naturaleza, ya que algunos de ellos desempeñan importantes papeles en la biosfera (por ejemplo la fijación del nitrógeno atmosférico por algunos microorganismos del suelo).
Es importante observar que no todas las células de un organismo pluricelular determinado son necesariamente de la misma clase: por ejemplo en las plantas superiores las células de las partes verdes (hojas y a veces tallos) son fotolitótrofas, mientras que las células de la raíz son quimioorganótrofas y dependen nutritivamente de los productos elaborados por las primeras. Es más, algunas células poseen una gran flexibilidad metabólica: las células de las hojas en las plantas superiores son fotolitótrofas durante el día y quimioorganótrofas durante la noche.
3.-RUTAS METABÓLICAS.
Los centenares de reacciones químicas que integran el metabolismo no tienen lugar de manera independiente unas de otras, sino que están articuladas en largas secuencias de reacciones consecutivas ligadas entre sí por intermediarios comunes, de manera que el producto de cada reacción resulta ser el sustrato o reactivo de la siguiente. Estas secuencias de reacciones reciben el nombre de rutas metabólicas.
La existencia de un intermediario común entre dos reacciones consecutivas hace posible la transferencia de energía química entre ellas. Por ejemplo en dos reacciones consecutivas tales como
A —› B
B —› C
parte de la energía química que reside en los enlaces de la sustancia A puede transferirse hasta la sustancia C a través del intermediario común B. Así, sobre la base de este principio del intermediario común, las rutas metabólicas constituyen eficaces medios para transferir la energía química desde aquellas reacciones exergónicas que la liberan hasta aquellas, endergónicas, que la requieren.
Las rutas metabólicas a su vez están organizadas en un complejo entramado en el que unas están conectadas con otras a través de encrucijadas metabólicas, en las cuales hay un metabolito común a dos o más rutas.
4.-FASES DEL METABOLISMO: CATABOLISMO Y ANABOLISMO.
El metabolismo se divide en dos fases principales: el catabolismo y el anabolismo.
El catabolismo es la fase degradativa del metabolismo, en la cual moléculas orgánicas complejas y relativamente grandes como los polisacáridos o las proteínas se degradan para dar lugar a moléculas de estructura más simple y menor tamaño tales como el ácido láctico, CO2, agua, amoníaco o urea. Este proceso degradativo va acompañado de la liberación de la energía química inherente a la estructura de las moléculas orgánicas que se degradan; es por lo tanto un proceso exergónico. Muchas reacciones del catabolismo suponen una oxidación, es decir, una pérdida de electrones, de los sustratos orgánicos que se degradan. En resumen, el catabolismo es un proceso degradativo, oxidante y exergónico.
En contrapartida, el anabolismo es la fase constructiva del metabolismo, en la cual tiene lugar la síntesis de los componentes moleculares de las células tales como los ácidos nucleicos, las proteínas, los polisacáridos y los lípidos a partir de moléculas precursoras de estructura más sencilla y menor tamaño. Este proceso biosintético requiere energía química para poder ser llevado a cabo, es decir, es un proceso endergónico. La construcción de biomoléculas orgánicas altamente hidrogenadas requiere electrones para reducir a sus precursores relativamente oxidados. En resumen, el anabolismo es un proceso constructivo, reductor y endergónico.
Las rutas metabólicas que forman parte del catabolismo se denominan rutas catabólicas, mientras que las que forman parte del anabolismo se denominan rutas anabólicas. Existen también algunas rutas que, en todo o en parte, son comunes al catabolismo y al anabolismo; reciben el nombre de rutas anfibólicas.
De lo expuesto anteriormente podría extraerse la falsa impresión de que catabolismo y anabolismo son procesos que transcurren por separado en el espacio y en el tiempo. En realidad ambos tienen lugar simultáneamente en el citoplasma celular puesto que las células están permanentemente en un proceso de renovación de sus componentes moleculares. Habría que considerar al catabolismo y al anabolismo, más que como fases, como dos "facetas" o "áreas de actividad" de una unidad funcional única que es el metabolismo.
5.-CONEXIONES ENERGÉTICAS EN EL METABOLISMO.
Como vimos en el apartado anterior el metabolismo incluye procesos que liberan energía (los procesos exergónicos del catabolismo) y otros que la consumen (los procesos endergónicos del anabolismo). Esta liberación y este consumo de energía no tienen por que ocurrir al mismo tiempo ni en el mismo lugar de la célula. Por lo tanto debe existir algún mecanismo que almacene esta energía y la transporte desde los lugares en que se libera hasta aquellos en que se consume, es decir, algún tipo de conexión energética entre el catabolismo y el anabolismo.
Dos son los sistemas que universalmente utilizan las células para llevar a cabo este almacenamiento y transporte de energía que conecta el catabolismo con el anabolismo: el sistema ADP/ATP y el sistema de los coenzimas transportadores de electrones.
5.1.-EL SISTEMA ADP/ATP.
Puesto que las células no pueden utilizar el calor como fuente de energía (son esencialmente isotermas), la energía que se desprende en los procesos exergónicos del catabolismo debe ser recuperada y almacenada en alguna otra forma más útil para producir trabajo, tal como la energía química inherente a ciertos enlaces.
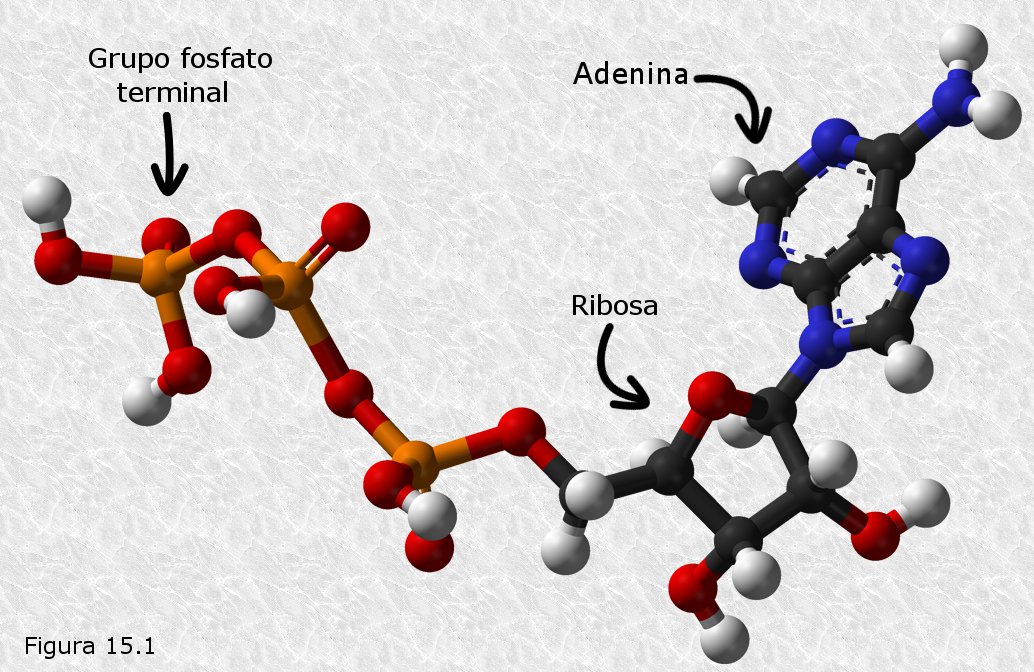
Las células recuperan y almacenan la energía desprendida durante los procesos degradativos del metabolismo en forma de la energía química del enlace fosfato terminal del trifosfato de adenosina (ATP) (Figura 15.1). La particular estructura química de este nucleótido hace que el enlace anhídrido que une sus grupos fosfato segundo y tercero sea un enlace rico en energía, es decir, un enlace que consume una cantidad importante de energía cuando se forma y que libera una cantidad importante de energía cuando se rompe.
La energía desprendida en las reacciones exergónicas del catabolismo se utiliza para formar enlaces fosfato terminales del ATP en un proceso endergónico que se denomina fosforilación y que tiene lugar mediante la reacción
ADP +Pi —› ATP +H2O
Existen dos mecanismos para acoplar el desprendimiento de energía durante el catabolismo con la síntesis de ATP:
a) Fosforilación a nivel de sustrato.- Se realiza en dos etapas. En la primera se forma un compuesto intermediario con algún enlace rico en energía. En la segunda se utiliza la energía desprendida en la hidrólisis de este compuesto para llevar a cabo la fosforilación. En el estudio de las distintas rutas catabólicas tendremos ocasión de ver varios ejemplos de este proceso.
b) Fosforilación acoplada al transporte electrónico.- El transporte de electrones a través de unas cadenas de transportadores ubicados en la membrana mitocondrial interna o en la membrana tilacoidal de los cloroplastos libera energía, la cual es utilizada por un enzima, la ATP-sintetasa, para fosforilar el ADP a ATP. Si este proceso tiene lugar en la mitocondria se denomina fosforilación oxidativa y si tiene lugar en el cloroplasto fosforilación fotosintética.
La energía así almacenada en forma de los enlaces fosfato terminales del ATP puede ahora ser ahora utilizada para impulsar las reacciones endergónicas del anabolismo mediante el acoplamiento de éstas con el proceso exergónico que es la hidrólisis del ATP:
ATP + H2O —› ADP + Pi
Este acoplamiento se realiza mediante enzimas que hacen posible la reacción global. Generalmente el ATP cede en primer lugar su grupo fosfato terminal al sustrato de la reacción para dar lugar a un intermediario fosforilado que a continuación se hidroliza para rendir fosfato inorgánico y el producto de la reacción.
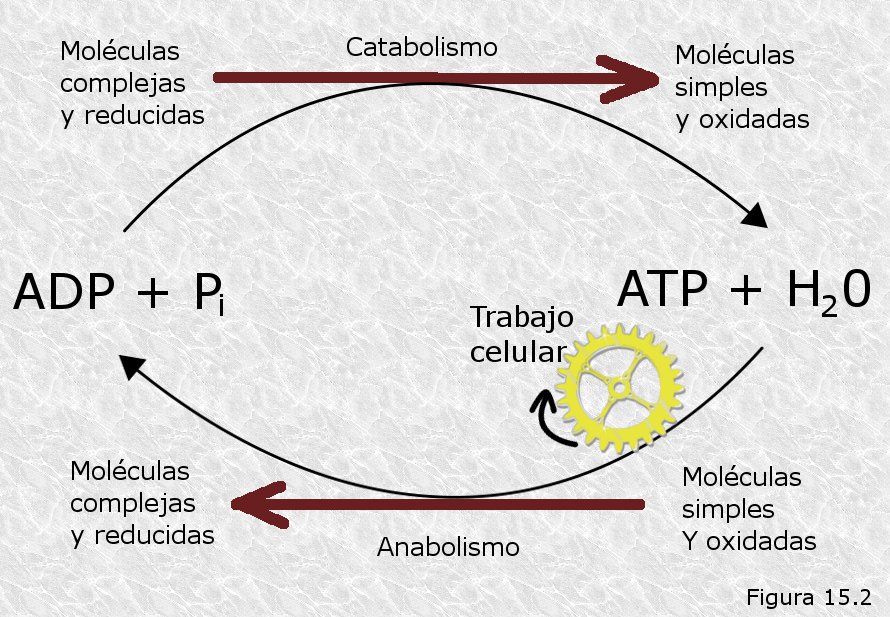
De lo dicho hasta aquí se deduce que el ATP viene a ser una especie de "moneda energética" de la célula ya que es la molécula que almacena y transporta la energía química desde los procesos que la liberan hasta los que la consumen (Figura 15.2). Aunque existen otros compuestos cuya hidrólisis libera mucha más energía que la del ATP, el "quántum" energético inherente a esta molécula parece ser el más adecuado para dosificar la energía de una manera eficaz, atendiendo así al principio de máxima economía que rige el metabolismo celular.
Por último, aunque el ATP es con mucho la molécula más utilizada por las células como almacén y transporte de energía, otros nucleótidos trifosfato pueden desempeñar funciones similares, como por ejemplo el UTP en la síntesis de polisacáridos o el GTP en la síntesis de proteínas. El ATP puede ceder su grupo fosfato terminal a diferentes nucleótidos difosfato para obtener los correspondientes nucleótidos trifosfato.
5.2.-COENZIMAS TRANSPORTADORES DE ELECTRONES.
Los electrones constituyen otro eficaz vehículo para canalizar hacia el anabolismo laenergía química desprendida en el catabolismo. Como sabemos, muchas reacciones del catabolismo suponen una oxidación de los sustratos orgánicos que se degradan, es decir, una pérdida de electrones por parte de éstos, mientras que la biosíntesis anabólica de moléculas orgánicas altamente hidrogenadas requiere electrones para reducir a sus precursores relativamente oxidados. Puesto que los procesos que liberan electrones y los que los requieren no tienen por que suceder simultáneamente ni en el mismo lugar de la célula, debe existir algún mecanismo para transportar dichos electrones entre estos dos tipos de proceso. Este mecanismo está integrado por una serie de coenzimas transportadores de electrones. Se trata de coenzimas cuya particular estructura química les permite aceptar o ceder electrones, es decir reducirse u oxidarse, de modo reversible.
Existen varios coenzimas transportadores de electrones. Químicamente todos son nucleótidos que poseen como parte de su estructura alguna de las bases nitrogenadas nicotinamida y flavina, en las cuales reside precisamente su capacidad para aceptar o ceder electrones. Estas bases nitrogenadas, que son diferentes a las que se encuentran habitualmente en los ácidos nucleicos, no pueden ser sintetizadas por la mayoría de los animales superiores, por lo que éstos deben incorporarlas en la dieta en forma de las vitaminas ácido nicotínico y riboflavina respectivamente. En la siguiente tabla se reflejan los coenzimas transportadores más importantes en sus formas oxidada y reducida. Las figuras 15.3 y 15.4 muestran las estructura química de los coenzimas más importantes
|
FORMA OXIDADA |
FORMA REDUCIDA |
|
NAD+ |
NADH + H+ |
|
NADP+ |
NADPH + H+ |
|
FAD |
FADH2 |
|
FMN |
FMN2 |
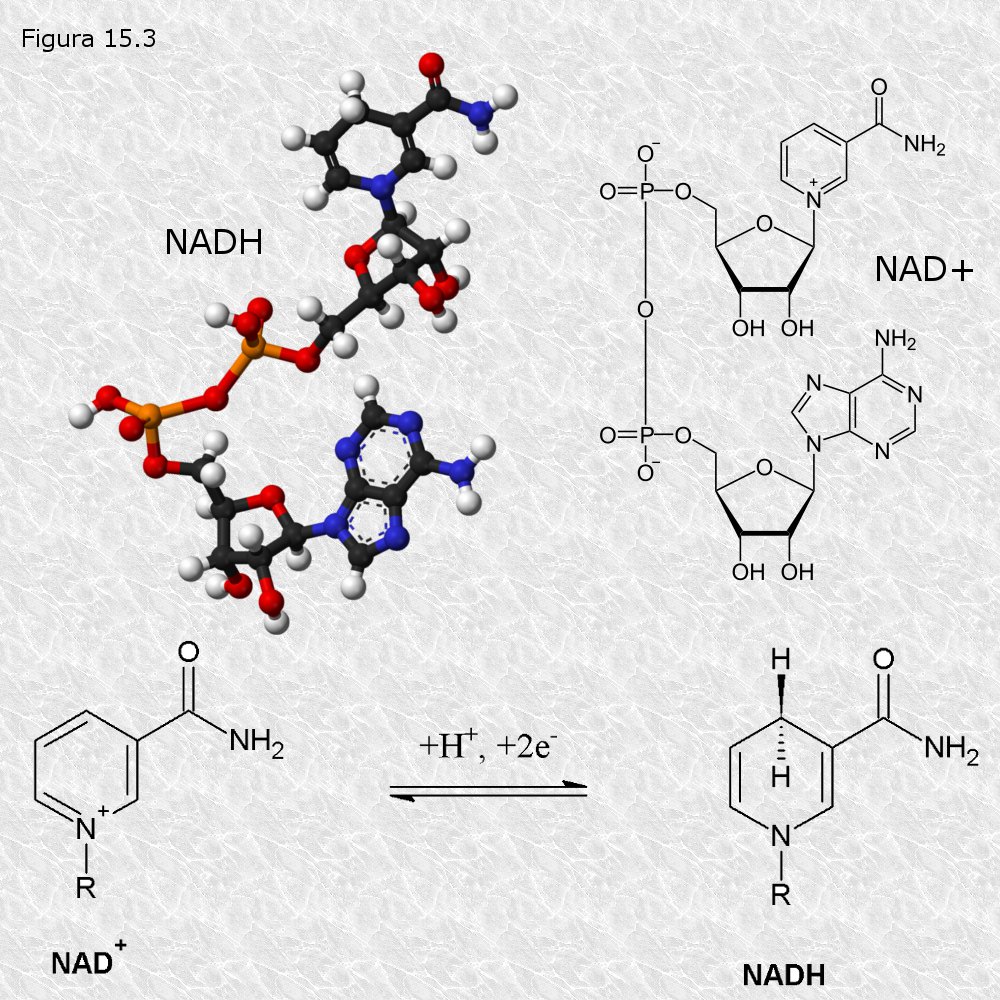
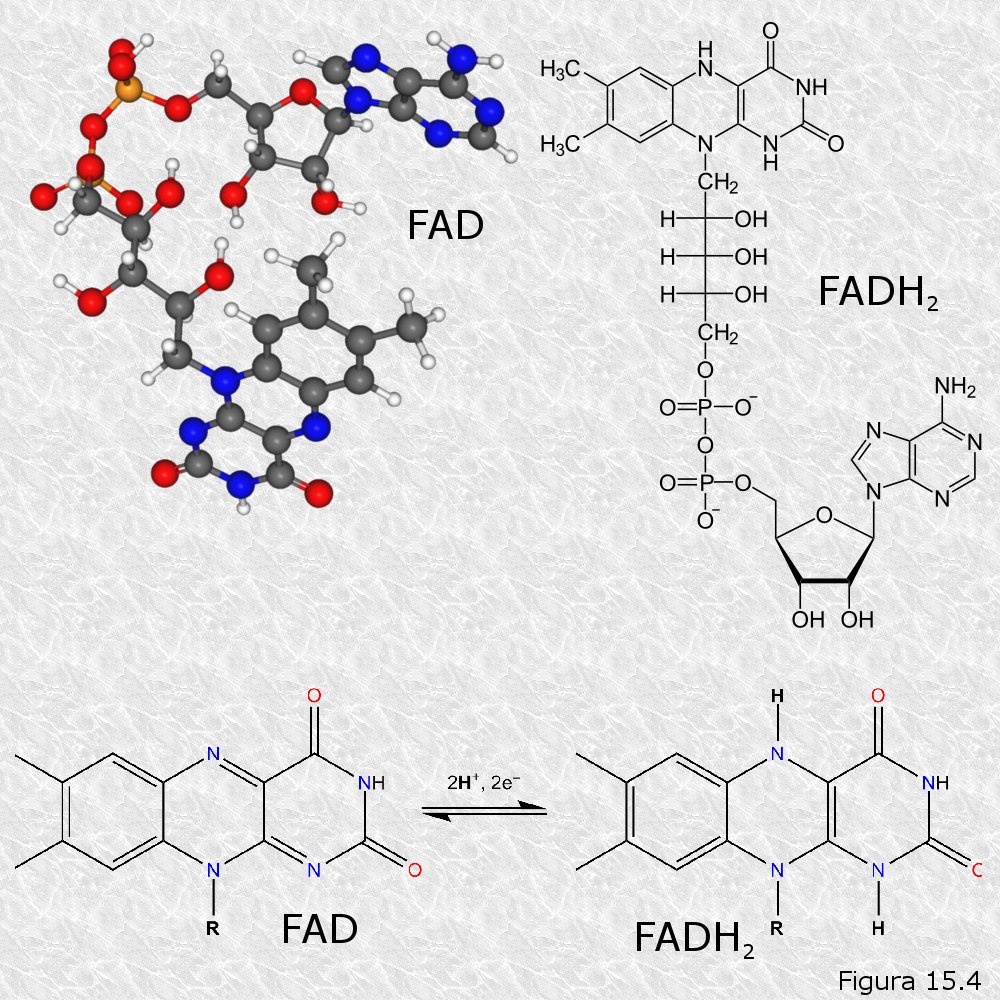
Se puede apreciar que los electrones siempre son aceptados o cedidos por pares, bien en forma de tales electrones o de átomos de hidrógeno, siendo acompañados en este último caso por los correspondientes protones.
No todos los electrones que se desprenden en las oxidaciones del catabolismo son canalizados directamente hacia las biosíntesis reductoras del anabolismo, sino que muchos de ellos son cedidos por los coenzimas transportadores a la cadena de transporte electrónico mitocondrial con el objeto de obtener ATP mediante el proceso de fosforilación oxidativa.
El estudio que hemos realizado sobre estos aspectos generales del metabolismo nos permitirá abordar con mayor garantía de comprensión el análisis detallado de las rutas del catabolismo y el anabolismo, cosa que haremos en los próximos temas