|
|
CURSO DE BIOLOGÍAAlejandro Porto Andión |
|
|
|
|
|
|
|
Inicio Temas de Biomoléculas Aula virtual |
|
|
|
|
![]()
TEMA 4: EL AGUA COMO BIOMOLÉCULA.
1.-INTRODUCCIÓN.
El agua es, con mucho, la sustancia más abundante en los sistemas vivos, constituyendo un 70% o más del peso de la mayoría de los organismos. Está presente en todos los lugares de la célula, es el medio de transporte de los nutrientes celulares y el medio de reacción en el que tiene lugar la inmensa mayoría de las reacciones químicas del metabolismo; es, en definitiva, el medio en el que se mueven e interactúan las demás biomoléculas.
La abundancia y ubicuidad del agua en la materia viva no deben conducirnos al error de considerarla como un líquido inerte con la única misión de rellenar espacios vacíos en los organismos vivos. Muy por el contrario, el agua participa activamente como reactivo en muchas reacciones químicas celulares y, lo que es más importante, la estructura y propiedades de muchas biomoléculas y otros componentes celulares dependen de su interacción con las moléculas de agua que los rodean.
2.- ESTRUCTURA MOLECULAR DEL AGUA: EL PUENTE DE HIDRÓGENO.
Como se puede comprobar en la tabla adjunta, el agua tiene un punto de fusión, punto de ebullición, calores de fusión y vaporización y tensión superficial más elevados que la mayoría de los líquidos comunes. Otras sustancias químicamente afines a ella, como el NH3 y el SH2, son gases a temperatura ambiente, mientras que el agua, como es obvio, se encuentra en estado líquido a esa temperatura. Estas propiedades físicas excepcionales delatan la existencia de poderosas fuerzas intermoleculares que mantienen unidas entre sí las moléculas de agua adyacentes, ya que de lo contrario, los cambios de estado de sólido a líquido y de líquido a gas no consumirían tanta energía como parecen indicar dichas propiedades. Analizaremos a continuación la estructura de la molécula de agua, con el objeto de identificar la naturaleza de esas fuerzas intermoleculares responsables de tan elevado grado de cohesión interna.
| Substancia | Punto de fusión (ºC) | Punto de ebullición (ºC) | Calor de vaporización (J/g) |
| AGUA | 0 | 100 | 2260 |
| METANOL | -98 | 65 | 1100 |
| ETANOL | -117 | 78 | 654 |
| PROPANOL | -127 | 97 | 687 |
| BUTANOL | -90 | 117 | 590 |
| ACETONA | -95 | 56 | 523 |
| HEXANO | -98 | 69 | 423 |
| BENCENO | 6 | 80 | 394 |
| BUTANO | -135 | -0,5 | 381 |
| CLOROFORMO | -63 | 61 | 247 |
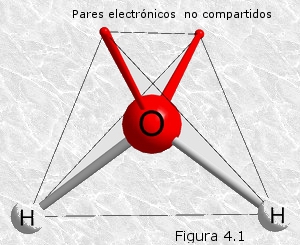
En la molécula de agua (H2O) cada átomo de hidrógeno comparte un par electrónico con el de oxígeno para formar un enlace covalente simple. La geometría de la molécula está determinada por la forma de los orbitales electrónicos más externos del átomo de oxígeno que son los que participan en los enlaces. Estos orbitales describen un tetraedro casi regular con los electrones compartidos con el hidrógeno en dos de sus vértices y los electrones sin compartir en los otros dos (Figura 4.1). De este modo, los tres átomos de la molécula forman un ángulo de 104,45º, algo menor que los 109,51º de un tetraedro regular, debido a que la repulsión entre los orbitales no enlazantes del oxígeno comprime ligeramente a los orbitales enlazantes con el hidrógeno.
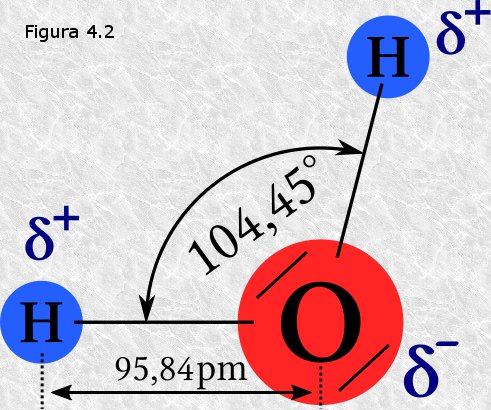
Por otra parte, el núcleo del átomo de oxígeno atrae hacia
sí con más fuerza los electrones compartidos que el núcleo del átomo de
hidrógeno, es decir, el átomo de oxígeno es más electronegativo.
Por lo tanto, en la molécula de agua el oxígeno y los hidrógenos
comparten los electrones de forma desigual: los electrones están más
tiempo cerca del oxígeno que del hidrógeno, de alguna manera
"pertenecen" más al oxígeno que al hidrógeno. Esta forma desigual de
compartir los electrones provoca la aparición de cargas eléctricas
parciales en distintas zonas de la molécula: sobre los átomos de
hidrógeno, parcialmente desposeídos de sus electrones, aparece, por
defecto, una carga parcial positiva (δ+);
correspondientemente el exceso de densidad electrónica sobre el átomo de
oxígeno se traduce en la aparición sobre el mismo de una carga parcial
negativa (δ-) que se encuentra repartida entre sus dos
orbitales no enlazantes
(Figura 4.2). Así, la molécula de agua, aún siendo
eléctricamente neutra (carga neta=0), se convierte en un dipolo
eléctrico doble, con una carga parcial positiva sobre cada átomo de
hidrógeno y una carga parcial negativa en cada uno de los orbitales no
enlazantes del oxígeno. Las cuatro cargas parciales se encuentran en
disposición tetraédrica alrededor del núcleo del oxígeno.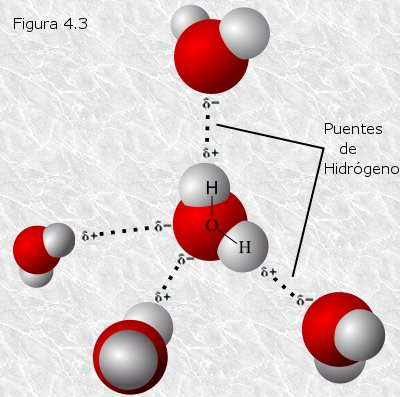
La existencia de cargas eléctricas parciales permite que se establezcan interacciones electrostáticas entre moléculas de agua vecinas: las cargas parciales de signo contrario situadas en una y otra molécula se atraen mutuamente. Este tipo particular de interacción electrostática se conoce con el nombre de puente de hidrógeno (Figura 4.3). Así, los puentes de hidrógeno entre moléculas de agua vecinas son las fuerzas intermoleculares responsables de la elevada cohesión interna del agua y, por lo tanto, de sus excepcionales propiedades físicas. Los cambios de estado en el agua implican la rotura de un cierto número de puentes de hidrógeno; los elevados puntos de fusión y ebullición, así como los elevados calores de fusión y vaporización, se explican porque es necesario emplear una cantidad considerable de energía para romper dichos puentes de hidrógeno.
Cada molécula de agua, con sus cuatro cargas parciales en
disposición tetraédrica, puede unirse mediante puentes de hidrógeno con
otras cuatro moléculas vecinas que a su vez se disponen tetraédricamente
alrededor de la molécula central (Figura
4.3). En el hielo, cada molécula de agua se encuentra unida de este
modo a exactamente cuatro de sus vecinas configurando una red cristalina
regular. Cuando el hielo se funde se rompen algunos puentes de hidrógeno
de manera que a temperatura ambiente cada molécula de agua está unida a
un promedio de 3,4 moléculas adyacentes. Si se compara la rigidez del
hielo con la extrema fluidez del agua líquida, resulta
 sorprendente esta
pequeña diferencia entre ambos en lo que se refiere al grado de ligazón
entre sus moléculas; si las moléculas del agua líquida están tan
intensamente unidas por puentes de hidrógeno ésta debería ser mucho más
viscosa. La explicación a este curioso fenómeno reside en la corta vida
del puente de hidrógeno. Dado que la energía implicada en la formación
de un puente de hidrógeno es del mismo orden de magnitud que la energía
térmica presente en el agua a temperatura ambiente, éstos se establecen
y se rompen con suma facilidad: se ha calculado que la vida media de un
puente de hidrógeno es del orden de 10-9 seg. Esta
circunstancia permite a las moléculas de agua una gran movilidad en un
radio corto, ya que cada una puede formar en muy poco tiempo sucesivos
puentes de hidrógeno con muchas de sus vecinas.
sorprendente esta
pequeña diferencia entre ambos en lo que se refiere al grado de ligazón
entre sus moléculas; si las moléculas del agua líquida están tan
intensamente unidas por puentes de hidrógeno ésta debería ser mucho más
viscosa. La explicación a este curioso fenómeno reside en la corta vida
del puente de hidrógeno. Dado que la energía implicada en la formación
de un puente de hidrógeno es del mismo orden de magnitud que la energía
térmica presente en el agua a temperatura ambiente, éstos se establecen
y se rompen con suma facilidad: se ha calculado que la vida media de un
puente de hidrógeno es del orden de 10-9 seg. Esta
circunstancia permite a las moléculas de agua una gran movilidad en un
radio corto, ya que cada una puede formar en muy poco tiempo sucesivos
puentes de hidrógeno con muchas de sus vecinas.
Por último, los puentes de hidrógeno no son exclusivos del agua. Se pueden formar fácilmente entre cualquier átomo electronegativo y un átomo de hidrógeno unido covalentemente a otro átomo electronegativo de la misma o de otra molécula. Puesto que en las biomoléculas abundan los grupos funcionales capaces de formar puentes de hidrógeno, este tipo de interacción tiene una gran importancia biológica. En la Figura 4.4 se puede apreciar un modelo espacial compacto de una molécula de agua.
3.-EL AGUA COMO DISOLVENTE.
Entre las excepcionales propiedades físicas del agua destaca por su importancia biológica la extraordinaria capacidad que presenta para disolver una amplia gama de sustancias. Otras propiedades son aprovechadas por algunos seres vivos en los que el agua desempeña funciones específicas, sin embargo, dado que la gran mayoría de las biomoléculas se encuentran en las células en disolución acuosa, las propiedades disolventes del agua son de importancia capital para todas las formas de vida.
Al igual que las demás propiedades físicas, la capacidad disolvente del agua está basada en su naturaleza dipolar, que le permite establecer interacciones electrostáticas con determinados tipos de solutos.
Podemos considerar tres tipos de sustancias en lo que se refiere a su solubilidad en agua: sustancias hidrofílicas, sustancias hidrofóbicas, y sustancias anfipáticas.
a) Sustancias hidrofílicas (del griego "amantes del agua").- Son netamente solubles en agua. Entre ellas podemos diferenciar las sustancias iónicas, que poseen carga eléctrica neta, y las sustancias polares, que presentan en su molécula cargas parciales. Muchas biomoléculas son sustancias iónicas, como las sales minerales y las biomoléculas orgánicas poseedoras de grupos funcionales ionizados al pH de la célula (por ejemplo los aminoácidos). Otras muchas son sustancias polares, como las biomoléculas orgánicas con grupos funcionales capaces de formar puentes de hidrógeno (por ejemplo los azúcares).
El agua es un buen disolvente de este tipo de sustancias porque su molécula, al presentar cargas parciales, puede establecer interacciones electrostáticas con las moléculas de soluto: cuando una sustancia iónica o polar se disuelve en agua las interacciones agua-soluto sustituyen de manera energéticamente favorable a las interacciones soluto-soluto de la red cristalina. En el caso de las sustancias polares estas interacciones son del tipo que conocemos con el nombre de puentes de hidrógeno. La siguiente animación ilustra este fenómeno.
La capacidad del agua para disolver sustancias hidrofílicas tiene su expresión matemática en su elevada constante dieléctrica. En la siguiente expresión
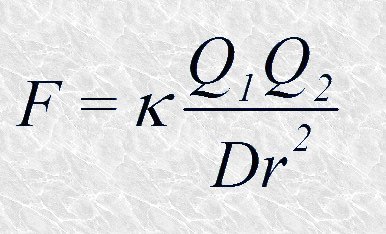
F es la fuerza con la que se atraen dos cargas eléctricas Q1 y Q2 separadas por una distancia r en un medio cuya constante dieléctrica es D. Cuanto mayor es el valor de D menor es la fuerza atractiva. Entre todos los líquidos conocidos el agua es el que tiene una mayor constante dieléctrica. Por lo tanto, cuando una sustancia iónica se encuentra en medio acuoso, las fuerzas electrostáticas que mantienen unidas sus moléculas en una red cristalina se debilitan considerablemente, lo que favorece en gran medida el proceso de disolución. En realidad, el elevado valor de la constante dieléctrica del agua no es más que una consecuencia de su carácter dipolar.
b) Sustancias hidrofóbicas (del griego "miedo al agua").- Son totalmente insolubles en agua. Se caracterizan por no poseer cargas eléctricas netas ni parciales, es decir, son totalmente apolares. Este carácter apolar les impide establecer interacciones energéticamente favorables con las moléculas de agua, es más, interfieren con los puentes de hidrógeno entre ellas, por lo que, cuando se encuentran en medio acuoso, tienden a agregarse y precipitar. De este modo ofrecen al agua la mínima superficie de contacto posible, y así se minimizan también las interferencias que ejercen sobre los puentes de hidrógeno entre sus moléculas. Algunas biomoléculas como las grasas neutras y las ceras son de naturaleza hidrofóbica; también lo son los gases biológicamente importantes, como el O2, el CO2 y el N2, que son muy poco solubles en agua.
c) Sustancias anfipáticas.-
Son sustancias que presentan en su molécula una parte polar (o cargada)
y otra no polar. Cuando estas sustancias se mezclan con el agua las dos
zonas de su molécula experimentan tendencias contrapuestas: la zonas
polares tienden a establecer interacciones electrostáticas con las
moléculas de agua mientras que las zonas no polares tienden a agregarse
para ofrecer la mínima superficie de contacto con ella. El resultado de
estas dos tendencias contrapuestas es que las moléculas anfipáticas se
asocian para constituir unas estructuras estables denominadas micelas
(Figura
4.5) en las que las zonas polares se disponen hacia el exterior, en
contacto con el agua, mientras que las zonas no polares lo hacen hacia
el interior, aisladas del contacto con el agua y unidas entre sí por
unas atracciones débiles llamadas interacciones hidrofóbicas. En
determinadas condiciones las sustancias anfipáticas en el seno del agua
pueden dar lugar a bicapas cerradas sobre sí mismas que
constituyen la base estructural de las membranas celulares. Algunas
biomoléculas importantes son sustancias anfipáticas; entre ellas se
encuentran los ácidos grasos, las proteínas globulares y una amplia
categoría de lípidos llamados lípidos de membrana.
polar. Cuando estas sustancias se mezclan con el agua las dos
zonas de su molécula experimentan tendencias contrapuestas: la zonas
polares tienden a establecer interacciones electrostáticas con las
moléculas de agua mientras que las zonas no polares tienden a agregarse
para ofrecer la mínima superficie de contacto con ella. El resultado de
estas dos tendencias contrapuestas es que las moléculas anfipáticas se
asocian para constituir unas estructuras estables denominadas micelas
(Figura
4.5) en las que las zonas polares se disponen hacia el exterior, en
contacto con el agua, mientras que las zonas no polares lo hacen hacia
el interior, aisladas del contacto con el agua y unidas entre sí por
unas atracciones débiles llamadas interacciones hidrofóbicas. En
determinadas condiciones las sustancias anfipáticas en el seno del agua
pueden dar lugar a bicapas cerradas sobre sí mismas que
constituyen la base estructural de las membranas celulares. Algunas
biomoléculas importantes son sustancias anfipáticas; entre ellas se
encuentran los ácidos grasos, las proteínas globulares y una amplia
categoría de lípidos llamados lípidos de membrana.
4.-DISOLUCIONES ACUOSAS EN LOS SERES VIVOS.
Puesto que el agua es el disolvente en el que se hallan disueltas la gran mayoría de las biomoléculas, resulta evidente que las propiedades de las disoluciones acuosas serán de una gran importancia para los seres vivos. Analizaremos a continuación qué tipos de disoluciones acuosas se hayan presentes en los seres vivos así como sus propiedades más relevantes desde el punto de vista biológico.
Se reconocen dos tipos de disoluciones acuosas de interés biológico atendiendo al tamaño de las partículas del soluto:
a) Disoluciones moleculares (o verdaderas).- En ellas las partículas del soluto miden menos de 10 nm. Cada partícula es una molécula individual.
b) Disoluciones coloidales.- En ellas el tamaño de las partículas de soluto oscila entre 10 nm y 100 nm. Estas partículas pueden ser agrupaciones de moléculas (por ejemplo micelas) o bien moléculas individuales de gran tamaño (macromoléculas).
Dado que las biomoléculas presentan tamaños muy variados podemos concebir el medio celular como una compleja disolución acuosa en la que coexisten multitud de solutos, algunos de tamaño molecular y otros muchos de tamaño coloidal.
La presencia de solutos disueltos altera la geometría característica de las agrupaciones de moléculas de agua. Cada molécula o ion del soluto interactúa con una serie de moléculas de agua a su alrededor obligándolas a ordenarse de manera diferente a como lo harían en ausencia de soluto. Esta alteración en la estructura del agua se manifiesta mediante la aparición de una serie de propiedades nuevas, características de la disolución, denominadas propiedades coligativas. Entre estas propiedades figura el descenso del punto de fusión, el aumento del punto de ebullición y la disminución de la presión de vapor, pero la que presenta un mayor interés biológico es la ósmosis, por lo que le prestaremos una especial atención.
Cuando dos disoluciones acuosas de diferente concentración como las representadas en la Figura 4.6 se encuentran separadas por una membrana semipermeable, es decir, por una membrana que permite el paso de moléculas de agua pero no de moléculas de soluto, la tendencia del disolvente a diluir al soluto se manifiesta mediante un flujo diferencial de agua a través de la membrana: pasa más agua desde la disolución más diluida a la más concentrada que en sentido inverso. Este fenómeno se conoce con el nombre de ósmosis, y su base físico-química consiste en que las interacciones electrostáticas entre las moléculas de agua y las de soluto, más abundantes en la disolución más concentrada, retienen en mayor medida a las moléculas de agua en el compartimento que alberga dicha disolución. El flujo diferencial al que hacemos referencia provoca un aumento del nivel del líquido en el compartimento de la disolución más concentrada. Cuando el líquido alcanza una determinada altura (h) la presión hidrostática generada por este volumen adicional de líquido contrarresta el mencionado flujo diferencial, alcanzándose el equilibrio cuando dicho flujo es de igual magnitud en ambos sentidos. Esta presión hidrostática, necesaria para alcanzar el equilibrio, se conoce con el nombre de presión osmótica.
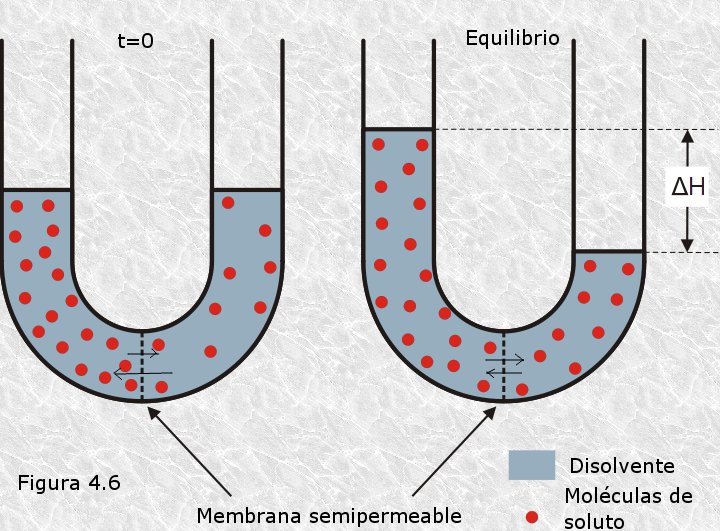
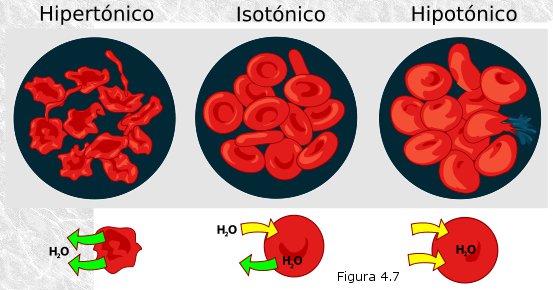
El interés biológico de esta propiedad de las disoluciones que llamamos ósmosis reside en que las membranas celulares son membranas semipermeables: permiten el libre paso de moléculas de agua pero ejercen una permeabilidad selectiva sobre la mayoría de las biomoléculas disueltas en ella. Como consecuencia, las células están sometidas a fenómenos osmóticos, que dependerán de la concentración de solutos en el medio en que se encuentran (Figura 4.7). Si la concentración de solutos en el medio es mayor que en el interior de la célula (medio hipertónico), ésta perderá agua por ósmosis sufriendo una retracción que en grado extremo acarreará la muerte celular. Si la concentración de solutos en el medio es igual a la del interior de la célula (medio isotónico), ésta estará en equilibrio osmótico con su entorno y no sufrirá cambios. Por último, si la concentración de solutos en el medio es inferior a la del interior celular (medio hipotónico), la consiguiente entrada de agua en la célula producirá un aumento de la presión osmótica en su interior. En un primer momento este aumento se traducirá en una mayor turgencia de la célula; más tarde, cuando la presión osmótica en el interior vence la resistencia mecánica de la membrana (que es muy limitada), sobrevendrá la lisis (rotura de la membrana con pérdida del contenido celular) y con ella la muerte de la célula.
Las células deben protegerse de alguna manera frente a los fenómenos osmóticos desfavorables a los que se hallan expuestas. Dos son las estrategias evolutivas que han adoptado para conseguirlo. La primera de ellas consiste en habitar exclusivamente en entornos isotónicos con respecto al interior celular, tal y como hacen algunos organismos unicelulares y también las células de los animales superiores, que se han dotado de un medio interno isotónico. La segunda, típica de las bacterias y de las células vegetales, consiste en dotarse de una pared celular resistente que les permite soportar las elevadas presiones osmóticas generadas por los medios hipotónicos en los que habitualmente viven.
Las disoluciones coloidales, además de presentar las propiedades características de todas las disoluciones, poseen otras adicionales como resultado del elevado tamaño de las partículas del soluto. Entre estas propiedades adicionales cabe destacar la sedimentación (capacidad de sedimentar las partículas del soluto por ultracentrifugación), la adsorción (tendencia de las partículas coloidales a adherirse a las superficies de determinados sólidos), la turbidez, etc. Algunas de estas propiedades de las disoluciones coloidales son de gran utilidad en los procesos de purificación y caracterización necesarios para el estudio de las macromoléculas.
5.- IONIZACIÓN DEL AGUA. ÁCIDOS Y BASES. TAMPONES.
Aunque gran parte de las propiedades del agua como disolvente se pueden explicar en función de su molécula sin carga (H2O), el pequeño grado de ionización del agua en iones hidrógeno e iones hidroxilo también debe ser tenido en cuenta. Además, los ácidos débiles y las bases débiles cuando se disuelven en agua alteran las cantidades relativas de estos iones en la disolución, bien aportando iones hidrógeno por disociación si son ácidos, o bien retirándolos por protonación si son bases. Por ello, dado que muchos procesos biológicos se ven afectados por las concentraciones de iones hidrógeno e hidroxilo, es conveniente que prestemos ahora atención a la ionización del agua y de los ácidos y bases débiles disueltos en ella.
La molécula de agua tiene una ligera tendencia a ionizarse reversiblemente dando lugar a un ion hidrógeno y un ion hidroxilo según la reacción
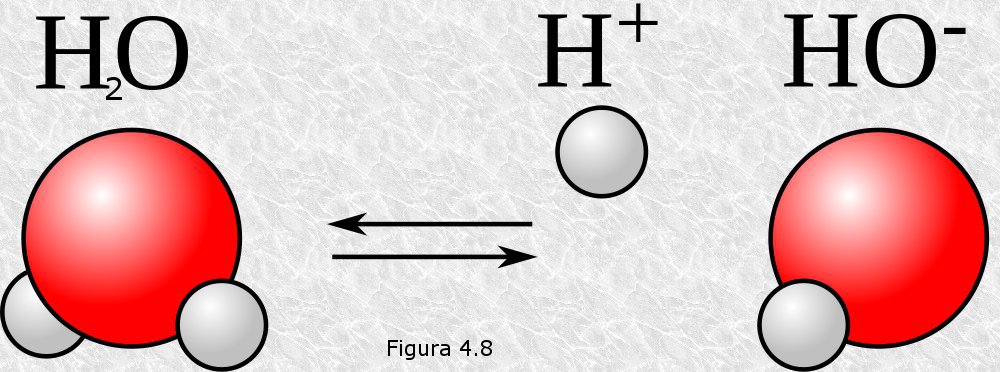
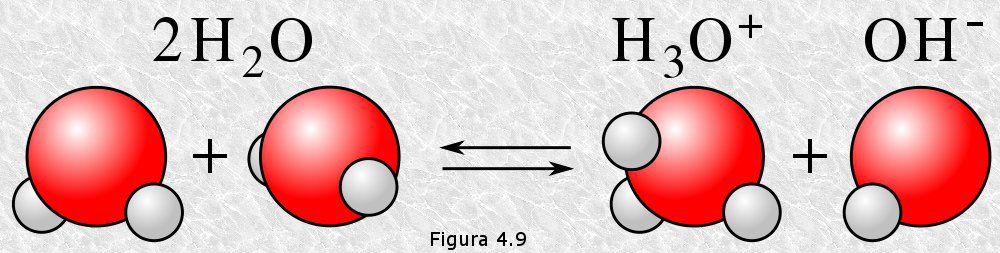
Aunque adoptaremos esta simplificación para analizar el proceso de autoionización del agua debemos tener en cuenta que este proceso no da en realidad lugar a iones hidrógeno "desnudos" sino a iones hidronio, como se puede apreciar en la figura 4.9
Para la ionización reversible del agua, como para cualquier otra reacción química, podemos escribir su constante de equilibrio:
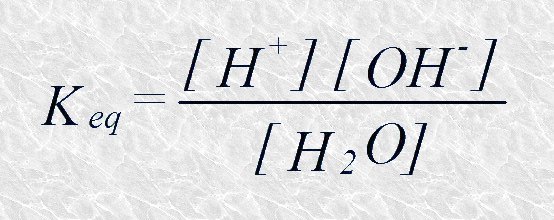
La constante de equilibrio de la ionización reversible del agua nos permite describir este proceso en términos cuantitativos, es decir, conocer en qué grado se encuentra ionizada el agua a una temperatura determinada.
El valor de [H2O] en el agua pura es 55,5 M (1.000 g de agua en un litro dividido por su masa molecular, o sea, 1.000/18=55,5). Dado que las concentraciones de iones hidrógeno e hidroxilo en el agua pura son muy bajas, este valor es esencialmente constante (no disminuye significativamente cuando el agua se ioniza), por lo que podemos sustituirlo en la ecuación anterior y reordenarla como sigue:
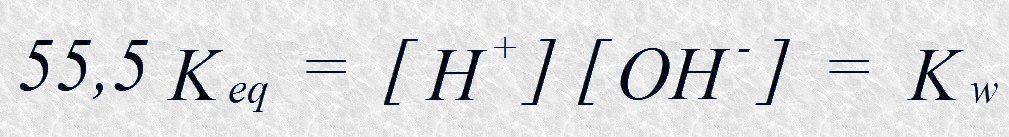
El término Kw se conoce como producto iónico del agua, y es constante para una temperatura dada. El valor de Keq, que se ha determinado a partir de las medidas de conductividad eléctrica en el agua pura, es de 1,8 x 10-16 M a 251C. Sustituyendo este valor en la ecuación anterior tenemos:
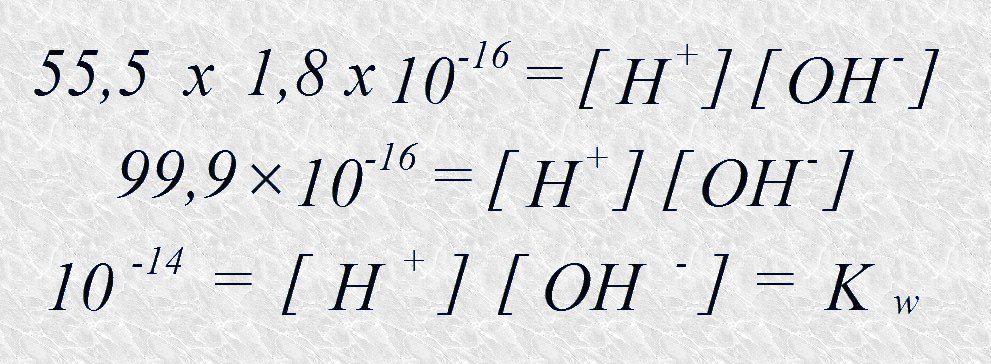
Así, el valor del producto iónico del agua a 25ºC es 10-14. A partir de este valor se puede calcular la concentración de iones hidrógeno e iones hidroxilo en el agua pura a 25ºC. Puesto que por cada molécula de agua que se disocia se obtiene un ion hidrógeno y un ion hidroxilo podemos deducir que
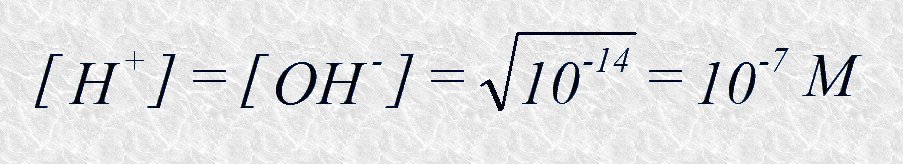
Es decir, en un instante dado sólo una de cada diez millones de moléculas se encuentra ionizada en el agua pura a 25ºC.
| [H+] (M) | pH | [OH-] (M) |
| 100 | 0 | 10-14 |
| 10-1 | 1 | 10-13 |
| 10-2 | 2 | 10-12 |
| 10-3 | 3 | 10-11 |
| 10-4 | 4 | 10-10 |
| 10-5 | 5 | 10-9 |
| 10-6 | 6 | 10-8 |
| 10-7 | 7 | 10-7 |
| 10-8 | 8 | 10-6 |
| 10-9 | 9 | 10-5 |
| 10-10 | 10 | 10-4 |
| 10-11 | 11 | 10-3 |
| 10-12 | 12 | 10-2 |
| 10-13 | 13 | 10-1 |
| 10-14 | 14 | 100 |
Tabla 4.2
En una disolución acuosa, si [H+] es igual a [OH-], tal como ocurre en el agua pura, se dice que la disolución es neutra; si [H+] es mayor que [OH-] se dice que es ácida; y si [H+] es menor que [OH-] se dice que es básica. En cualquier caso, el producto de ambas concentraciones (Kw) es constante e igual a 10-14 a 25ºC, de tal modo que si una de las dos concentraciones es elevada la otra debe ser proporcionalmente reducida para que el producto iónico del agua permanezca constante (Tabla 4.2).
A partir del producto iónico del agua se introduce la escala de pH. La escala de pH constituye una forma ágil y cómoda de expresar las concentraciones de iones hidrógeno (y consiguientemente de iones hidroxilo) en disoluciones acuosas diluidas. Su uso evita tener que manipular cifras engorrosas del tipo de 0,000001 M o 10-8 M. Es válida para expresar concentraciones entre 10-14 M y 1 M de iones hidrógeno (ver Tabla 4.2). El pH se define como el logaritmo negativo de la concentración de iones hidrógeno:
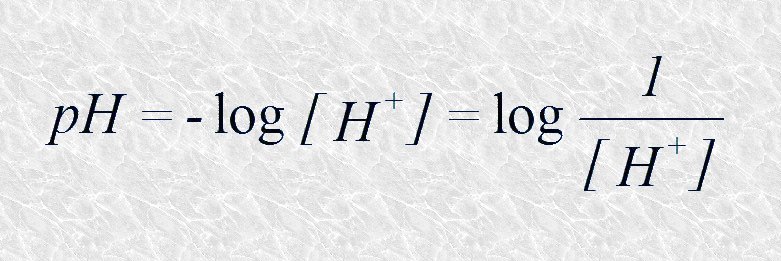
Una disolución acuosa neutra tiene un pH=7 ya que en ella [H+]=10-7 y por lo tanto:
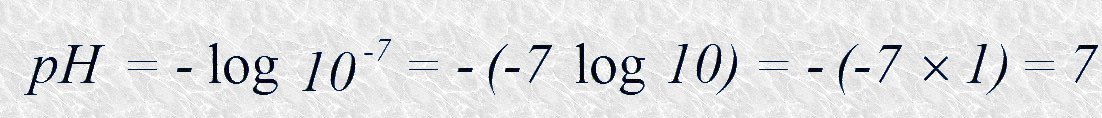
De la misma manera, el pH de una disolución ácida será menor que 7 y el de una disolución básica mayor que 7. Obsérvese que la escala de pH es logarítmica, no aritmética: decir que dos disoluciones difieren en una unidad de pH significa que una de ellas posee una concentración de iones hidrógeno diez veces superior a la de la otra.
El pH de una disolución puede medirse de manera aproximada utilizando indicadores (sustancias que cambian de color con las variaciones de pH), o de una manera más precisa mediante aparatos denominados pHmetros. La medida del pH es una de las operaciones más corrientes en la investigación bioquímica ya que sus variaciones tienen efectos importantes sobre muchas biomoléculas.
Aunque existen definiciones de ácidos y bases de aplicación más general, los conceptos introducidos en su día por Brönsted y Lowry son de mayor utilidad para describir el comportamiento de ácidos y bases en sistemas acuosos como los que estamos tratando. Según el concepto de Brönsted-Lowry un ácido es un dador de protones, y una base es un aceptor de protones. A cada ácido corresponde una base conjugada de modo que, aplicando este formulismo, ácidos y bases no se tratan por separado, sino como integrantes de pares ácido-base conjugados. Por ejemplo, el ácido acético, un dador de protones, y el anión acetato, su correspondiente aceptor, forman un par ácido-base conjugado relacionado por la reacción reversible
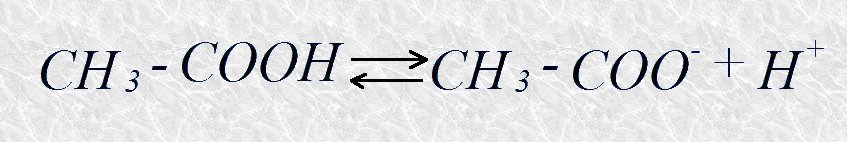
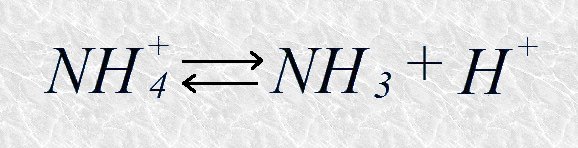
El amoníaco, un aceptor de protones, es la base conjugada del ion amonio, su correspondiente dador, según la reacción
Cada ácido tiene una tendencia característica a ceder su protón. Los ácidos fuertes tienen una tendencia elevada a ceder su protón, mientras que los ácidos débiles lo ceden con más dificultad. A un ácido fuerte corresponde una base conjugada débil (con poca afinidad por el protón) mientras que a un ácido débil corresponde una base conjugada fuerte (con gran afinidad por el protón). La mayor o menor tendencia de un ácido AH a ceder su protón para formar la base conjugada A- según la reacción
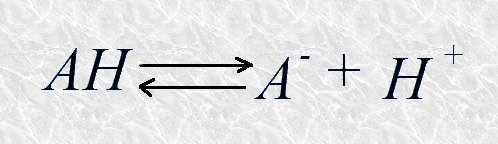
puede expresarse de un modo cuantitativo mediante la constante de equilibrio de dicha reacción, que en este caso recibe el nombre de constante de disociación (Ka)

Los ácidos fuertes presentan valores altos de Ka, mientras que los ácidos débiles presentan valores bajos de Ka. Estas constantes de disociación son características de cada par conjugado ácido-básico para una temperatura determinada.
Al igual que ocurre con los valores de [H+] en las disoluciones acuosas diluidas, los valores de Ka resultan a veces engorrosos en lo que a su manipulación se refiere. Por ello, de manera análoga a como se introdujo el concepto de pH, introduciremos ahora el concepto de pK, que se define como el logaritmo negativo de la constante de disociación:
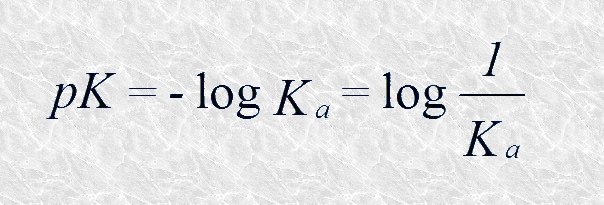
Cuanto más fuerte sea un ácido más bajo será el valor de su pK; por el contrario, los ácidos débiles presentan pK's elevados. Debemos tener en cuenta que el pK expresa tanto la tendencia de un ácido a ceder protones como la de su base conjugada a aceptarlos; es, pues, una constante característica de cada par conjugado ácido-básico a una temperatura determinada.
La titulación es un procedimiento que se usa habitualmente en química para conocer la concentración de un ácido en una disolución. Se parte de un volumen conocido de la disolución problema a la que se va añadiendo gota a gota otra disolución de una base fuerte (generalmente NaOH) de concentración conocida. Cuando se produce la neutralización, lo que se determina mediante un colorante indicador, se puede determinar la concentración inicial del ácido a partir del volumen de disolución básica añadido y de su concentración. Si en el curso de una titulación vamos midiendo periódicamente con un pHmetro el pH de la disolución problema, y lo representamos gráficamente frente a la cantidad de disolución básica añadida, obtendremos la curva de titulación característica del par ácido-básico titulado. En la Figura 4.10 se representa a modo de ejemplo la curva de titulación del ácido acético (HAc). Esta curva revela cómo va variando el pH y las cantidades relativas de la especie dadora y aceptora de protones a medida que transcurre el proceso de titulación. En este proceso están implicados dos equilibrios químicos cuyas constantes características deben permanecer inalteradas a lo largo de toda la titulación:
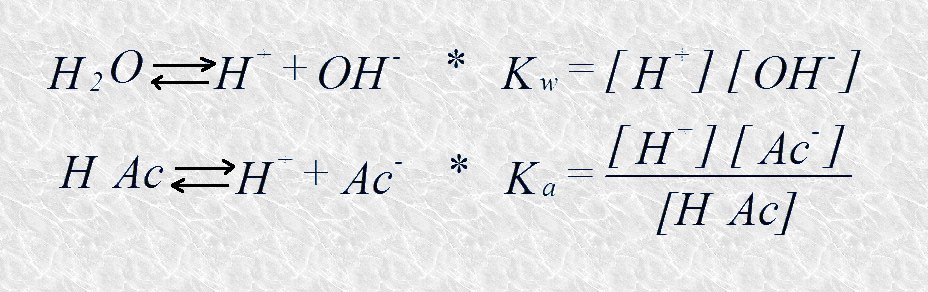
Al principio, antes de añadir hidróxido sódico, el ácido acético está sólo ligeramente disociado, en un grado que viene dado por su constante de disociación; a medida que se va añadiendo hidróxido sódico, los iones hidroxilo adicionados se van combinando con los iones H+ libres en la disolución para formar agua de modo que el producto iónico de ésta permanezca constante; al ir desapareciendo iones H+ libres el ácido acético se disocia para formar acetato y liberar nuevos iones H+, satisfaciendo así su propia constante de disociación; al final del proceso todo el ácido acético inicial habrá cedido sus protones transformándose en acetato.
Las curvas de titulación de diferentes pares ácido-base conjugados presentan siempre la misma forma y sólo se diferencian en su posición más o menos desplazada verticalmente sobre la escala del pH (Figura 4.11). Este hecho sugiere que dichas curvas deben reflejar algún tipo de relación matemática común a todas ellas. Tal relación se conoce con el nombre de ecuación de Henderson-Hasselbalch, y se puede deducir fácilmente reenunciando de manera conveniente la expresión de la constante de disociación de un ácido débil (Ka) tal y como sigue:
Despejando [H+] tenemos:
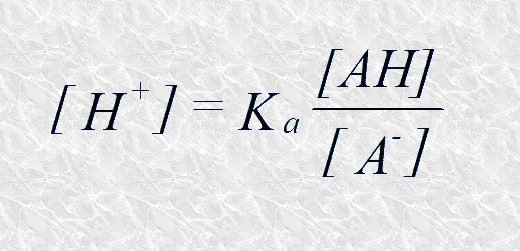
Tomando ahora el logaritmo negativo de ambos miembros:
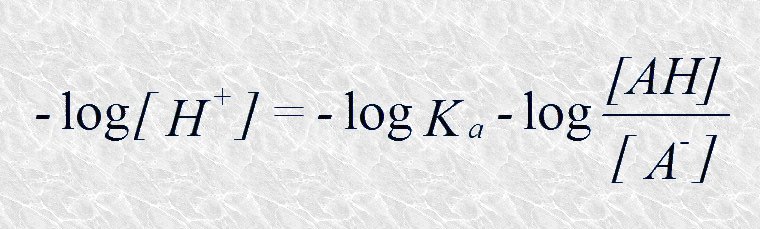
Sustituyendo -log [H+] por pH y -log Ka por pK tenemos:
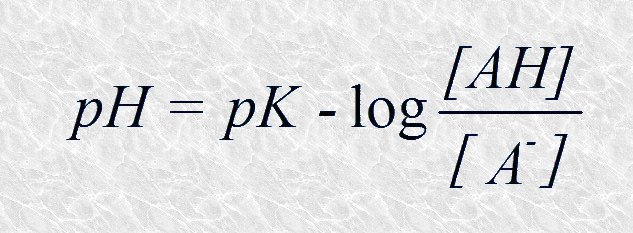
Y cambiando el signo:
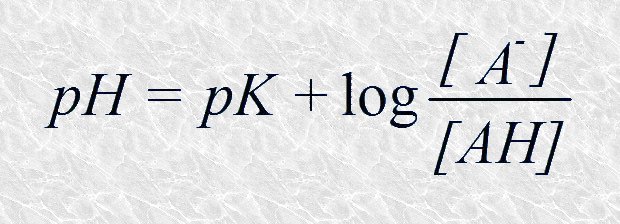
Esta última expresión es la ecuación de Henderson-Hasselbalch que describe la forma de la curva de titulación de cualquier par conjugado ácido-básico en el que AH es la especie dadora de protones y A- la especie aceptora de protones. La ecuación de Henderson-Hasselbalch es de gran utilidad en el tratamiento cuantitativo de los sistemas ácido-base pues permite, por ejemplo, calcular el pH de una disolución conociendo el pK del par conjugado y la relación molar entre la especie aceptora y la especie dadora de protones, o bien calcular el pK del par conjugado a partir del pH y de dicha relación molar. Sin embargo, el principal interés biológico de esta ecuación, y de las curvas de titulación que describe, es que ponen de manifiesto cómo un par conjugado ácido-básico puede funcionar como sistema tampón. Nos ocuparemos de ello seguidamente.
Los sistemas tampón son disoluciones acuosas que tienden a responder a la adición de cantidades moderadas de ácidos o de bases con pequeñas variaciones en su pH. Ilustraremos su funcionamiento a través del siguiente ejemplo: si se añade 1 ml de HCl 10 N a un litro de agua pura (pH 7) el pH descenderá aproximadamente hasta un valor de 2; si se añade la misma cantidad de HCl a un litro de plasma sanguíneo, que tiene su propio sistema tampón, el pH sólo descenderá desde 7,4 hasta aproximadamente 7,2. Un sistema tampón (o simplemente un tampón), consiste en una disolución de un par conjugado ácido-básico que presenta concentraciones aproximadamente iguales de la especie dadora y de la especie aceptora de protones. Si se añade un ácido a una disolución de tales características, lo que implica un aumento en la concentración de iones H+, el par conjugado reacciona desplazando su equilibrio de disociación en el sentido de retirar iones H+ de la disolución de manera que el pH no varía o lo hace muy poco. Si por el contrario se añade una base, los iones hidroxilo retirarán iones hidrógeno de la disolución para formar agua y el par conjugado reaccionará desplazando su equilibrio de disociación en el sentido de liberar más iones H+ consiguiendo así que el pH tampoco varíe. Así pues, el efecto tampón es el resultado de la acción conjunta de dos equilibrios químicos: el de ionización del agua y el de disociación del par conjugado ácido-base. Tanto la especie dadora como la aceptora de protones tienen que estar presentes en la disolución para que el par conjugado funcione como tampón.
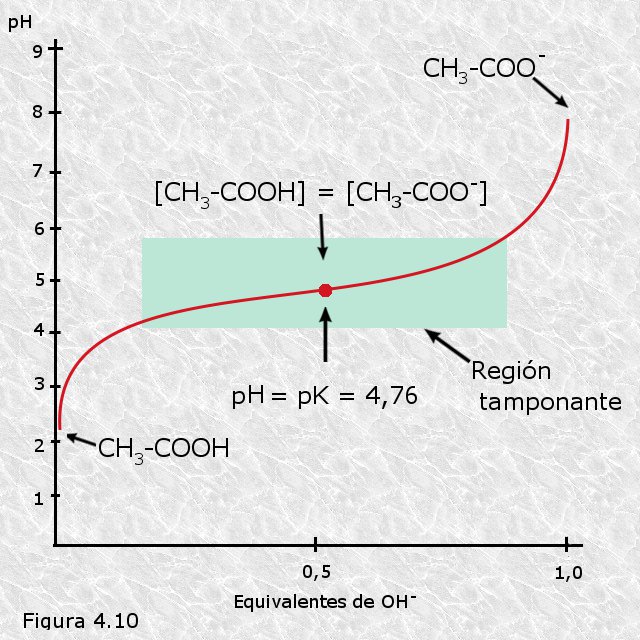
Las curvas de titulación de los diferentes pares conjugados ácido-básicos reflejan perfectamente este efecto tampón: todas ellas presentan un tramo casi horizontal a lo largo del cual las variaciones de pH son mínimas en relación a la cantidad de disolución básica añadida (ver Figuras 4.10 y 4.11). En el punto medio de esta región tamponante, exactamente cuando se igualan las concentraciones de la especie dadora y de la especie aceptora de protones, la capacidad de tamponamiento del par conjugado es máxima. Aplicando la ecuación de Henderson-Hasselbalch podemos deducir que en este punto el pH de la disolución es numéricamente igual al pK del par conjugado:
Puesto que en el punto medio de la región tamponante [A-]=[AH]:
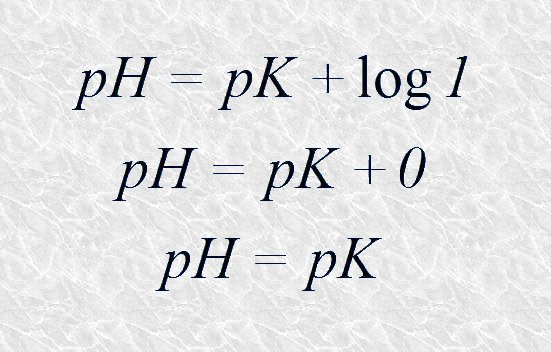
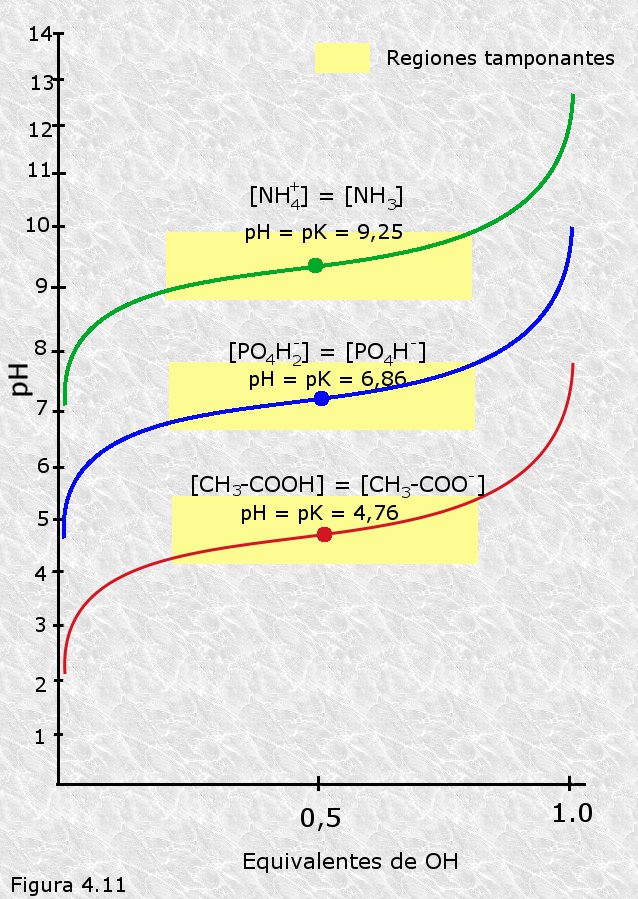 Por lo que podemos afirmar que todo par conjugado ácido-básico tiene
capacidad de tamponamiento para valores de pH próximos a su pK. La
región tamponante propia de cada par conjugado se extiende
aproximadamente 0,5 unidades de pH a ambos lados su punto medio, en el
que se igualan los valores de pH y pK. Más allá de estos límites el par
conjugado no tiene capacidad de tamponamiento.
Por lo que podemos afirmar que todo par conjugado ácido-básico tiene
capacidad de tamponamiento para valores de pH próximos a su pK. La
región tamponante propia de cada par conjugado se extiende
aproximadamente 0,5 unidades de pH a ambos lados su punto medio, en el
que se igualan los valores de pH y pK. Más allá de estos límites el par
conjugado no tiene capacidad de tamponamiento.
Los tampones tienen una gran importancia biológica ya que muchos procesos vitales se ven influidos de una u otra manera por las variaciones en el pH del medio acuoso en el que transcurren. Entre las biomoléculas abundan las que contienen grupos funcionales ionizables como los grupos amino y carboxilo; del grado de ionización de dichos grupos funcionales, que se ve claramente afectado por los cambios en el pH del medio, depende en muchos casos la actividad biológica de la molécula. Por ejemplo, los enzimas, de cuya actividad depende en gran medida la vida de las células, tienen un ph óptimo para el cual su actividad catalítica es máxima, y se inactivan rápidamente si el pH del entorno varía más allá de unos límites estrechos con respecto a dicho pH óptimo. Por esta razón, los seres vivos se ven obligados a regular el pH de sus células y fluidos corporales de manera que el estado iónico de sus biomoléculas sea el más adecuado para que éstas puedan desempeñar correctamente sus funciones. Para ello utilizan tampones, que mantienen constante el pH de estos fluidos en unos valores que, en la mayor parte de los casos, están próximos a la neutralidad (pH 7).
Muchas biomoléculas poseen grupos funcionales ionizables que contribuyen al poder tamponante de los fluidos biológicos; sin embargo, existen dos sistemas tampón que, por estar presentes en muchas formas de vida, tienen una especial relevancia: el sistema tampón del fosfato, que regula el pH en el citoplasma de todas las células, y el sistema tampón del bicarbonato, que actúa en la sangre y en los líquidos intersticiales de los vertebrados. Estos son sus equilibrios de disociación:
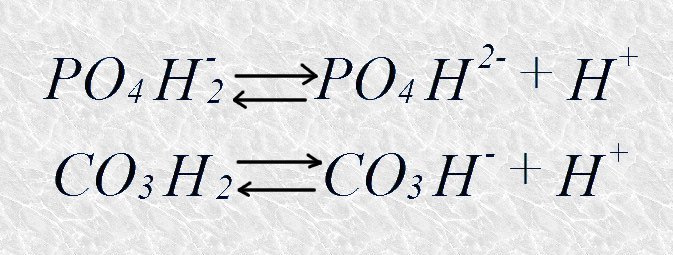
El sistema tampón del fosfato, con un pK de 6,86, presenta clara capacidad de tamponamiento al pH celular, que está próximo a la neutralidad. Resulta curioso sin embargo, que el sistema tampón del bicarbonato, con un pK de 3,77, tenga capacidad de tamponamiento para valores de pH próximos a 7. Esta capacidad reside en que el ácido carbónico, aún estando casi totalmente disociado a estos valores de pH, puede reponerse a partir del CO2 disuelto en la sangre, que a su vez está en equilibrio con el CO2 gaseoso presente en los pulmones.
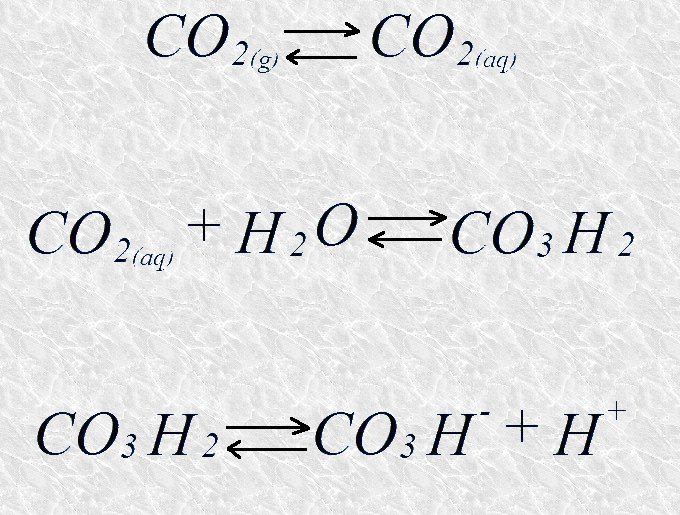
La acción conjunta de estos tres equilibrios químicos es la responsable del poder tamponante a pH fisiológico del tampón bicarbonato.
6.-INTERACCIONES DÉBILES EN LOS SISTEMAS ACUOSOS.
Como ya se explicó en el capítulo precedente, las llamadas interacciones débiles entre átomos o grupos de átomos vecinos son de una extraordinaria importancia biológica ya que de ellas dependen muchos de los procesos en los que descansa el fenómeno de la vida. Las circunstancias en que tienen lugar estas interacciones, así como la mayor o menor fortaleza de las mismas, se ven influidas de manera determinante por la naturaleza acuosa del medio en el que tienen lugar.
Las interacciones débiles se caracterizan porque las cantidades de energía que se ponen en juego en su formación o ruptura (energías de enlace) son netamente inferiores a las correspondientes a los verdaderos enlaces químicos: la energía necesaria para romper un enlace covalente de tipo medio es del orden de 100 kcal/mol, mientras que las energías necesarias para romper los diferentes tipos de interacciones débiles oscilan entre 1 y 5 kcal/mol.
Consideraremos cuatro tipos de interacciones débiles de importancia biológica: los puentes de hidrógeno, las interacciones iónicas, las interacciones hidrofóbicas y las fuerzas de Van der Waals.
a) Puentes de hidrógeno.- Como ya hemos visto, son interacciones electrostáticas entre cargas parciales, que aparecen cuando un átomo de hidrógeno unido covalentemente a un átomo electronegativo se halla próximo a otro átomo electronegativo. En las biomoléculas existe una gran variedad de grupos funcionales capaces de establecer puentes de hidrógeno entre sí. Estos puentes pueden ser intramoleculares, si se dan entre grupos funcionales presentes en una misma molécula, o intermoleculares, si dichos grupos se encuentran en moléculas diferentes.
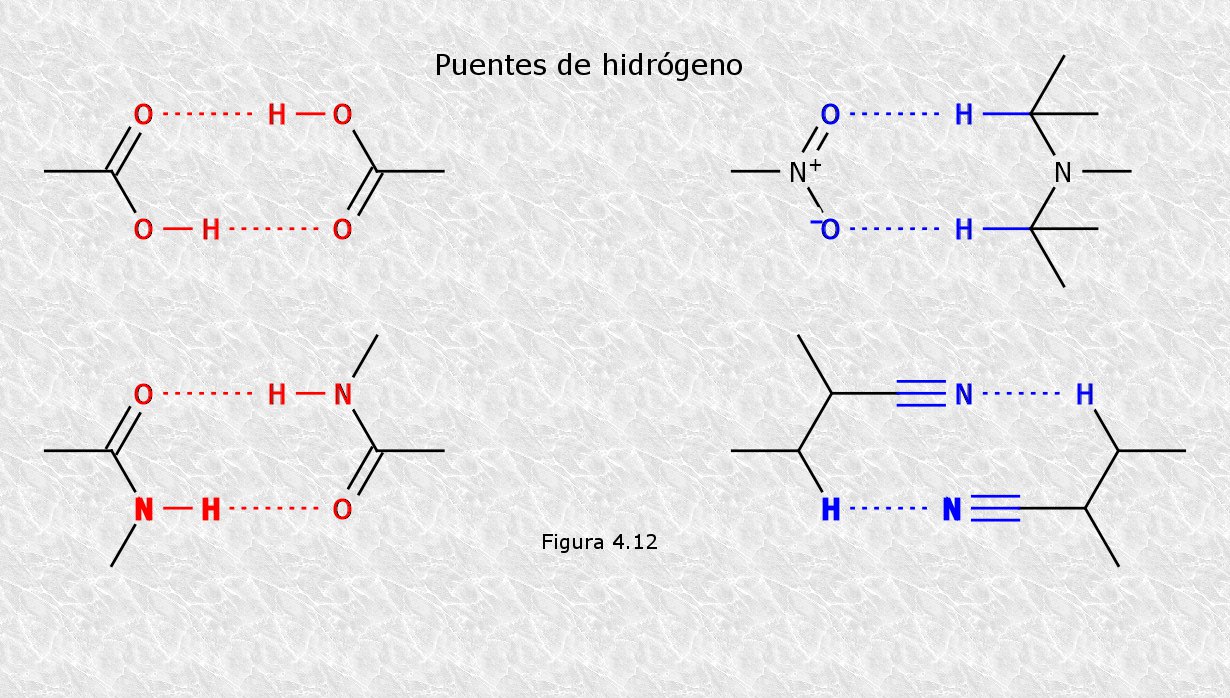
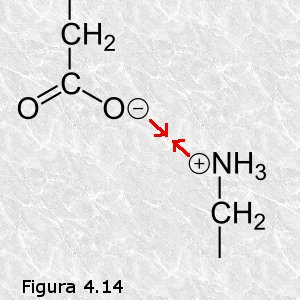
b) Interacciones iónicas.- Son interacciones electrostáticas entre átomos o grupos de átomos que presentan carga eléctrica neta. Pueden ser atractivas o repulsivas, siendo aquellas las de mayor importancia biológica. En los diferentes tipos de biomoléculas existen grupos funcionales que presentan carga neta a pH 7 y que por lo tanto son susceptibles de participar en este tipo de interacciones (Figura 4.14).
La razón de que incluyamos a las interacciones iónicas de importancia biológica entre las llamadas débiles y no entre los verdaderos enlaces químicos estriba en que, por tener lugar en medio acuoso, estas interacciones tienen energías de enlace sensiblemente inferiores a las que tendrían de tener lugar en cualquier otro medio. Ello es debido a que el agua, gracias a su elevada constante dieléctrica, tiene el efecto de reducir considerablemente las fuerzas electrostáticas que operan en su seno.
c) Interacciones hidrofóbicas.- Se producen cuando zonas hidrofóbicas de una misma molécula o de moléculas diferentes se aproximan entre sí, excluyendo el agua que las rodeaba, para establecer entre ellas un contacto estable, tal y como ocurre, por ejemplo, en las micelas. La fuerza de estas interacciones no reside en una atracción intrínseca entre las regiones hidrofóbicas, sino en la tendencia de éstas a ofrecer al agua la mínima superficie de contacto posible (Figura 4.13).

Las interacciones hidrofóbicas intervienen en procesos como la determinación de la estructura tridimensional de las proteínas o la construcción de las membranas celulares por lo que son de una gran importancia biológica.
d) Fuerzas de Van der Waals.- Cuando átomos no cargados se encuentran muy próximos entre sí, las variaciones aleatorias en la posición de los electrones alrededor de los núcleos pueden inducir la aparición sobre dichos átomos de dipolos eléctricos transitorios. Las atracciones electrostáticas entre estos dipolos se denominan fuerzas de Van der Waals. Son fuerzas muy débiles, inespecíficas y de naturaleza fluctuante, sin embargo, la acción combinada de muchas de ellas puede conferir gran estabilidad a una asociación molecular dada.
En capítulos sucesivos iremos viendo numerosos ejemplos que pondrán de manifiesto la gran importancia biológica de las interacciones débiles.
7.-EL AGUA COMO REACTIVO.
El agua no es tan sólo el disolvente en el que tienen lugar las reacciones químicas de las células vivas, sino que a menudo participa efectivamente como reactivo en dichas reacciones. Un tipo de reacción química muy común en los seres vivos es la hidrólisis (del griego "rotura por el agua") en la que una molécula de agua se adiciona a un enlace químico rompiéndolo. Esta reacción participa en la degradación biológica de distintos tipos de biomoléculas.
En otros casos, el agua es un producto de las reacciones químicas del metabolismo celular. Por ejemplo, en el proceso de la respiración celular distintos tipos de biomoléculas energéticas se oxidan en presencia de oxígeno para rendir CO2 y H2O. El "agua metabólica" así formada es de hecho suficiente para permitir que algunos animales que viven en ambientes muy secos puedan sobrevivir sin beber agua durante largos períodos.
8.-OTRAS FUNCIONES DEL AGUA EN LOS SERES VIVOS.
Además de ser el disolvente en el que se mueven e interactúan las demás biomoléculas, lo que constituye su principal función biológica, el agua desempeña otras funciones importantes en los seres vivos, que se han adaptado eficazmente a su entorno acuoso desarrollando medios para explotar en su provecho las excepcionales propiedades físicas del agua. Así, el elevado calor específico del agua es aprovechado por los animales homeotermos para regular su temperatura corporal. Además, algunos vertebrados utilizan el elevado calor de vaporización del agua para eliminar el exceso de calor mediante la evaporación del sudor. Por otra parte, el alto grado de cohesión interna del agua líquida da lugar a fenómenos de capilaridad que son aprovechados por las plantas para transportar los nutrientes en ella disueltos desde la raíz hasta las hojas. Incluso el que el agua líquida sea más densa que el hielo tiene consecuencias importantes para los organismos acuáticos: los estanques, lagos y mares se congelan en invierno desde arriba hacia abajo de manera que la capa superficial de hielo aísla al agua subyacente del aire frío impidiendo así que ésta se congele.
La influencia del agua en la evolución biológica ha sido profunda y determinante. Los seres vivos se han ido adaptando progresivamente al entorno acuoso en el cual se originaron. Si han aparecido formas de vida en algún otro lugar del universo es razonable pensar que en ese lugar deben existir grandes cantidades de agua disponible como disolvente.